OUTLINE CLASSIFICATION:
All living organisms are classified into groups based on very basic, shared characteristics. Organisms within each group are then further divided into smaller groups. These smaller groups are based on more detailed similarities within each larger group. This grouping system makes it easier for scientists to study certain groups of organisms. Characteristics such as appearance, reproduction, mobility, and functionality are just a few ways in which living organisms are grouped together. These specialized groups are collectively called the classification of living things. The classification of living things includes 7 levels: kingdom, phylum, classes, order, families, genus, and species.
1. BACTERIA AND ARCHAEA
The organisms most commonly found in such extremely acidic environments belong to the two major groups of prokaryotes: Bacteria and Archaea. The bacteria live in almost every environment on Earth. The archaea are a superficially similar group of microscopic, unicellular
prokaryotes. However, both the biochemistry and the genetics of bacteria differ in numerous ways from those of archaea. Not until the 1970s did biologists discover how radically different bacteria and archaea really are. And only with the sequencing of an archaeal genome in 1996 did we realize just how extensively archaea differ from both bacteria and eukaryotes. Many biologists acknowledge the antiquity of these clades and the importance of their differences by recognizing three domains of living things: Bacteria, Archaea, and Eukarya. The domain Bacteria comprises the “true bacteria.” The domain Archaea (Greek archaios, “ancient”) comprises other prokaryotes once called (inaccurately) “ancient bacteria.” The domain Eukarya includes all other living things on Earth. Dividing the living world in this way, with two prokaryotic domains and a single domain for all the eukaryotes, fits with the current trend toward reflecting evolutionary
relationships in classification systems. This section focuses on the two prokaryotic domains.
Why Three Domains?
What does it mean to be different? You and the person nearest you look very different-certainly you appear more different than the two cells. But the two of you are members of the same species, while these two tiny organisms that look so much alike actually are classified
in entirely separate domains. Still, all three of you (you in the domain Eukarya and those two prokaryotes in the domains Bacteria and Archaea) have a lot in common.
Members of all three domains
- conduct glycolysis;
- replicate their DNA semiconservatively;
- have DNA that encodes polypeptides;
- produce these polypeptides by transcription and
translation and use the same genetic code; - have plasma membranes and ribosomes in abundance.
There are also major differences among the domains. Members of the Eukarya have cells with nuclei, membraneenclosed organelles, and a cytoskeleton- structures that no prokaryote has. And a glance at Table 1 will show you that there are also major differences even under the
(most of which cannot be seen microscope) between the two prokaryotic domains. In some ways the archaea are more like us; in other ways they are more like bacteria.
Genetic studies have led many biologists to conclude that all three domains had a single common ancestor, and that the present-day archaea share a more recent Common ancestor with eukaryotes than they do with bacteria (Figure 1). Because of the ancient time at which
these three clades diverged, the major differences among the three kinds of organisms, and especially the likelihood that the archaea are more closely related to the eukaryotes than are either of those groups to the bacteria, many biologists agree that. it makes sense to
treat these three groups as domains-a higher taxonomic category than kingdoms. To treat all the prokaryotes as a single kingdom within a five-kingdom classification of organisms would result in a kingdom that is paraphyletic. That is, a single kingdom “Prokaryotes” would not include all the descendants of their common ancestor.
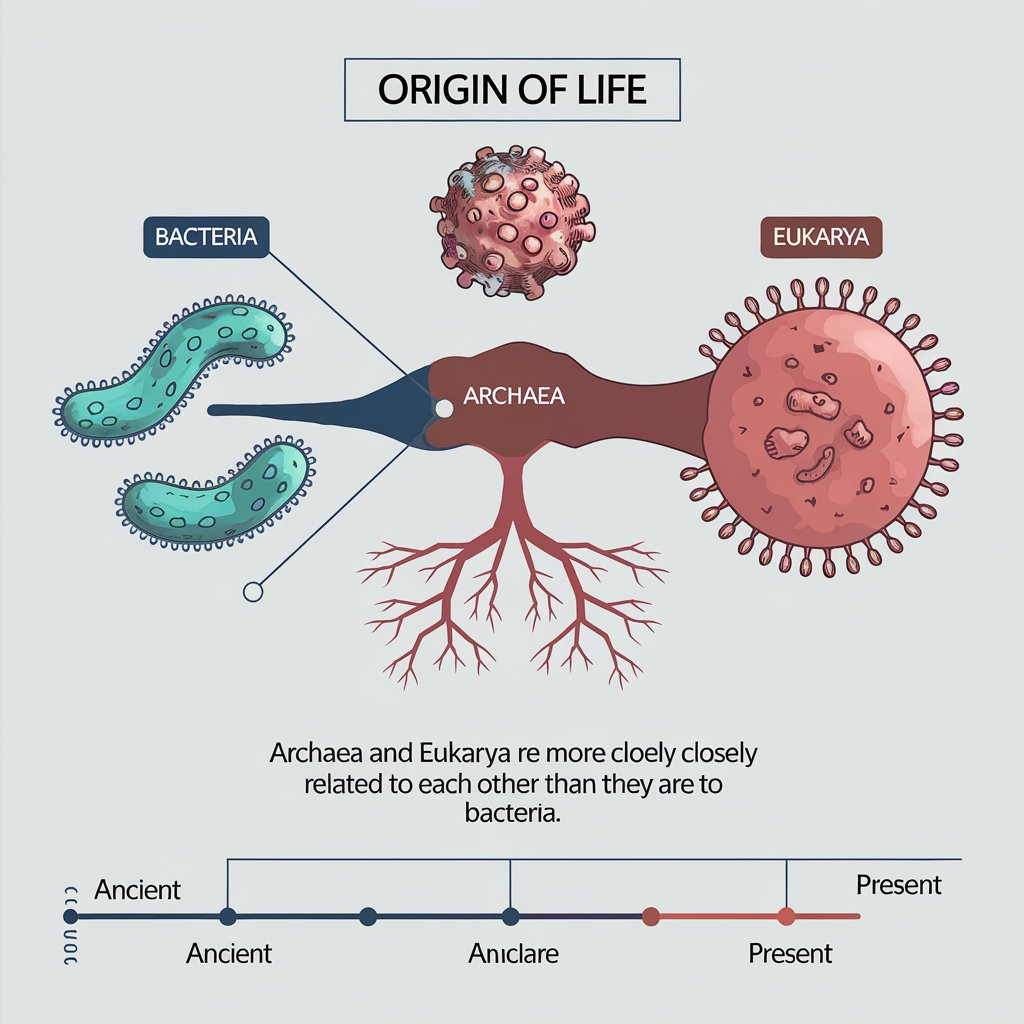
The common ancestor of all three domains was prokaryotic. Its genetic material was DNA; its machinery for trancription and translation produced RNAs and proteins, respectively. It probably had a circular chromosome, and many of its structural genes were grouped into operons.
The Archaea, Bacteria, and Eukarya of today are all products of billions of years of natural selection and genetic drift, and they are all well adapted to present-day environments. None are “primitive.” The common ancestor of the Archaea and the Eukarya probably lived more than 2 billion years ago, and the common ancestor of the Archaea, the Eukarya, and the Bacteria probably lived more than 3 billion years ago. The earliest prokaryotic fossils date back at least 3.5 billion years, and these ancient fossils indicate that there was considerable diversity among the prokaryotes even during the earliest days of life. The prokaryotes were alone on Earth for a very long time, adapting to new environments and to changes in existing environments. They have survived to this day-and in massive numbers.
General Biology of the Prokaryotes
There are many, many prokaryotes around us- everywhere. Although most are so small that we cannot see them with the naked eye, the prokaryotes are the most successful of all creatures on Earth, if success is measured by numbers of individuals. The bacteria in one person’s intestinal tract, for example, outnumber all the humans who have ever lived, and even the total number of human cells in their host’s body. Some of these bacteria form a thick lining along the intestinal wall. Bacteria and archaea in the oceans number more than 3 x 1028. This stunning number is perhaps 100 million times as great as the number of stars in the visible universe. Although small, prokaryotes play many critical roles in the biosphere, interacting in one way or another with every other living thing. In this section, we’ll see that some prokaryotes perform key steps in the cycling of nitrogen, sulfur, and carbon. Other prokaryotes trap energy from the sun or from inorganic chemical sources, and some help animals digest their food. The members of the two prokaryotic domains outdo all other groups in metabolic diversity. Eukaryotes, in contrast, are much more diverse in size and shape, but their metabolism is much less diverse. In fact, much of the energy metabolism of eukaryotes is carried out in organelles mitochondria and chloroplasts-that are descended from bacteria.
Prokaryotes are found in every conceivable habitat on the planet, from the coldest to the hottest, from the most acidic to the most alkaline, and to the saltiest. Some live where oxygen is abundant and others where there is no oxygen at all. They have established themselves at the bottom of the seas, in rocks more than 2 km into Earth’s solid crust, and inside other organisms, large and small. Their effects on our environment are diverse and profound. What do these tiny but widespread organisms look like?
Prokaryotes and their associations take a few characteristic forms
Three shapes are particularly common among the prokaryotes: spheres, rods, and curved or spiral forms. A spherical prokaryote is called a coccus (plural, cocci). Cocci may live singly or may associate in two- or three- dimensional arrays as chains, plates, blocks, or clusters
of cells. A rodshaped prokaryote is called a bacillus (plural, bacilli). Spiral forms are the third main prokaryotic shape. Bacilli and spiral forms may be single or may form chains. Prokaryotes are almost all unicellular, although some multicellular ones are known. Associations such as chains do not signify multicellularity because each cell is fully viable and independent. These associations arise as cells adhere to one another after reproducing by fission. Associations in the form of chains are called filaments.
Prokaryotes lack nuclei, organelles, and a cytoskeleton
The basic unit of archaea and bacteria is the prokaryotic cell, which contains a full complement of genetic and protein-synthesizing systems, including DNA, RNA, and all the enzymes needed to transcribe and translate the genetic information into proteins. The prokaryotic cell also contains at least one system for generating the ATP it needs.
In what follows, bear in mind that most of what we know about the structure of prokaryotes comes from studies of bacteria. We still know relatively little about the diversity of archaea, although the pace of research on archaea is accelerating. The prokaryotic cell differs from the eukaryotic cell in three important ways. First, the organization and replication of the genetic material differs. The DNA of the prokaryotic cell is not organized within a membrane-enclosed nucleus. DNA molecules in prokaryotes (both bacteria and archaea) are usually circular; in the best-studied prokaryotes, there is a single chromosome, but there are often plasmids as well.
Second, prokaryotes have none of the membrane- enclosed cytoplasmic organelles that modern eukaryotes have-mitochondria, Golgi apparatus, and others.
However, the cytoplasm of a prokaryotic cell may containa variety of infoldings of the plasma membrane and photosynthetic membrane systems not found in eukaryotes. Third, prokaryotic cells lack a cytoskeleton, and, without the cytoskeletal proteins, they lack mitosis.
Prokaryotic cells divide by their own elaborate method, fission, after replicating their DNA.
Prokaryotes have distinctive modes of locomotion
Although many prokaryotes cannot move, others are motile. These organisms move by one of several means.
Some spiral bacteria, called spirochetes, use a corkscrew- like motion made possible by modified flagella, called axial filaments, running along the axis of the cell beneath the outer membrane. Many cyanobacteria and a few other bacteria use various poorly understood gliding
mechanisms, including rolling. Various aquatic prokaryotes, including some cyanobacteria, can move slowly up and down in the water by adjusting the amount of gas in gas vesicles. By far the most common type of locomotion in prokaryotes, however, is that driven by flagella.

Bacterial flagella are slender filaments that extend singly or in tufts from one or both ends of the cell or are randomly distributed all around it .
A bacterial flagellum consists of a single fibril made of the protein flagellin, projecting from the cell surface, plus a hook and basal body responsible for motion. In contrast, the flagellum of eukaryotes is enclosed by the plasma membrane and usually contains a circle of nine pairs of microtubules surrounding two central microtubules, all containing the protein tubulin, along with many other associated proteins. The prokaryotic flagellum rotates about its base, rather than beating as a eukaryotic flagellum or cilium does.
Prokaryotes have distinctive cell walls
Most prokaryotes have a thick and relatively stiff cell wall. This wall is quite different from the cell walls of plants and algae, which contain cellulose and other polysaccharides, and from those of fungi, which contain chitin. Almost all bacteria have cell walls containing peptidoglycan (a polymer of amino sugars). Archaeal cell walls are of differing types, but most contain significant amounts of protein. One group of archaea has pseudopeptidoglycan in its wall; as you have probably already guessed from the prefix pseudo-, pseudopeptidoglycan is similar to, but distinct from, the peptidoglycan of bacteria. Peptidoglycan is a substance unique to bacteria; its absence from the walls of archaea is a key difference between the two prokaryotic domains.
In 1884 Hans Christian Gram, a Danish physician, developed a simple staining process that has lasted into our high technology era as a useful tool for identifying bacteria.
The Gram stain separates most types of bacteria into two distinct groups, Gram-positive and Gram-negative, on the basis of their staining. A smear of cells on a microscope slide is soaked in a violet dye and treated with iodine; it is then washed with alcohol and counterstained with safranine (a red dye). Gram- positive bacteria retain the violet dye and appear blue to purple. The alcohol washes the violet stain out of Gram-negative cells; these cells then pick up the safranine counterstain and appear pink to red. Gram-staining characteristics are useful in classifying some kinds of
bacteria and are important in determining the identity of bacteria in an unknown sample.
For many bacteria, the Gram-staining results correlate roughly with the structure of the cell wall. Peptidoglycan forms a thick layer outside the plasma membrane of Gram- positive bacteria. The Gram-negative cell wall usually has only one-fifth as much peptidoglycan, and outside the peptidoglycan layer the cell is surrounded by a second, outer membrane quite distinct in chemical makeup from the plasma membrane. Between the inner (plasma) and outer membranes of Gram-negative bacteria is the periplasmic space. This space contains enzymes that are important in digesting some materials, transporting others, and detecting chemical gradients in the environment.
The consequences of the different features of prokaryotic cell walls are numerous and relate to the disease-causing characteristics of some prokaryotes. Indeed, the cell wall is a favorite target in medical combat against diseases
that are caused by prokaryotes because it has no counterpart in eukaryotic cells. Antibiotics such as penicillin and ampicillin, as well as other agents that specifically interfere with the synthesis of peptidoglycan- containing cell walls, tend to have little, if any, effect on the cells of humans and other eukaryotes.



Prokaryotes reproduce asexually, but genetic recombination does occur
Prokaryotes reproduce by fission, an asexual process. Recall, however, that there are also processes- transformation, conjugation, and transduction-that allow the exchange of genetic information between some prokaryotes quite apart from either sex or reproduction.
Some prokaryotes multiply very rapidly. One of the fastest is the bacterium Escherichia coli, which under optimal conditions has a generation time of about 20 minutes. The shortest known prokaryote generation times are about 10 minutes. Generation times of 1 to 3 hours are common for others; some extend to days. Bacteria living deep in Earth’s crust may suspend their growth for more than a century without dividing and then multiply for a few days before suspending growth again. What kinds of metabolism support such a diversity of growth rates?
Prokaryotes have exploited many metabolic possibilities
The long evolutionary history of the bacteria and archaea, during which they have had time to explore a wide variety of habitats, has led to the extraordinary diversity of their metabolic “lifestyles”-their use or nonuse of oxygen, their energy sources, their sources of carbon atoms, and the materials they release as waste products.
ANAEROBIC VERSUS AEROBIC METABOLISM
Some prokaryotes can live only by anaerobic metabolism because molecular oxygen is poisonous to them. These oxygen-sensitive organisms are called obligate anaerobes.
Other prokaryotes can shift their metabolism between anaerobic and aerobic modes and thus are called facultative anaerobes. Many facultative anaerobes alternate between anaerobic metabolism (such as fermentation) and cellular respiration as conditions dictate. Aerotolerant anaerobes cannot conduct cellular respiration, but are not damaged by oxygen when it is present.
At the other extreme from the obligate anaerobes, some prokaryotes are obligate aerobes, unable to survive for extended periods in the absence of oxygen. They require oxygen for cellular respiration.
NUTRITIONAL CATEGORIES
Biologists recognize four broad nutritional categories of organisms: photoautotrophs, photoheterotrophs, chemolithotrophs, and chemoheterotrophs. Prokaryotes are represented in all four groups,
2 How Organisms Obtain Their Energy and Carbon
| Nutritional Category | Energy Source | Carbon Source |
| Photoaubtotrophs (found in all three domains) | light | carbon dioxide |
| Photoheterotrophs (some bacteria) | light | Organic compounds |
| Chemolithotrophs (some bacteria, many archaea) |
Inorganic substances | carbon dioxide |
| Chemoheterotrophs (found in all three domains) | Organic compounds | Organic compounds |
Photoautotrophs perform photosynthesis. They use light as their source of energy and carbon dioxide as their source of carbon. Like the photosynthetic eukaryotes, one group of photoautotrophic bacteria, the cyanobacteria, use chlorophyll a as their key photosynthetic pigment and produce oxygen as a by-product of noncyclic electron transport.
By contrast, the other photosynthetic bacteria use bacteriochlorophyll as their key photosynthetic pigment, and they do not release oxygen gas. Some of these photosynthesizers produce particles of pure sulfur instead because hydrogen sulfide (H2S), rather than H,0, is their electron donor for photophosphorylation. Bacteriochlorophyl l absorbs light of longer wavelengths than the chlorophyll used by all other photosynthesizing organisms does. As a result, bacteria using this pigment can grow in water beneath fairly dense layers of algae,0using light of wavelengths that are not absorbed by the algae.
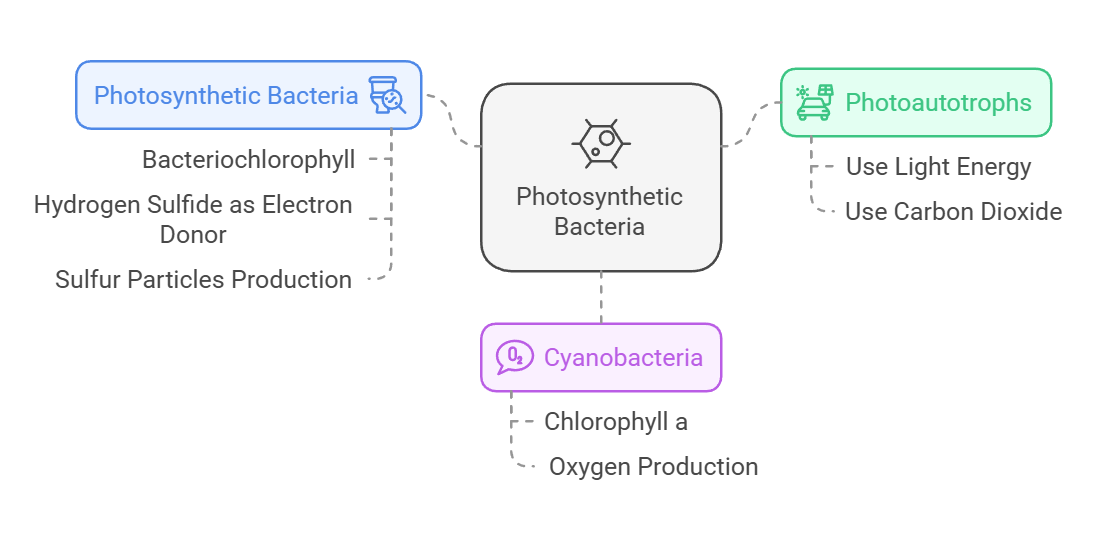
Photoheterotrophs use light as their source of energy, but must obtain their carbon atoms from organic compounds made by other organisms. They use compounds such as carbohydrates, fatty acids, and alcohols as their organic “food.” The purple nonsulfur bacteria, among others, are photoheterotrophs Chemolithotrophs (chemoautotrophs) obtain their energy by oxidizing inorganic substances, and they use some of that energy to fix carbon dioxide. Some chemolithotrophs use reactions identical to those of the typical photosynthetic cycle, but others use other pathways to fix carbon dioxide.
Some bacteria oxidize ammonia or nitrite ions to form nitrate ions. Others oxidize hydrogen gas, hydrogen sulfide, sulfur, and other materials. Many archaea are chemolithotrophs. Deep-sea hydrothermal vent ecosystems are based on chemolithotrophic prokaryotes
that are incorporated into large communities of crabs, mollusks, and giant worms, all living at a depth of 2,500 meters, below any hint of light from the sun. These bacteria obtain energy by oxidizing hydrogen sulfide and other substances released in the near-boiling water that
flows from volcanic vents in the ocean floor.
Finally, chemoheterotrophs obtain both energy and carbon atoms from one or more complex organic compounds. Most known bacteria and archaea are chemoheterotrophs- as are all animals and fungi and many protists.
NITROGEN AND SULFUR METABOLISM
Many prokaryotes base important parts of their metabolism on reactions involving nitrogen or sulfur. For example, some bacteria carry out respiratory electron transport without using oxygen as an electron acceptor. These organisms use oxidized inorganic ions such as nitrate, nitrite, or sulfate as electron acceptors. Examples include the denitrifiers, bacteria that release nitrogen to the atmosphere as nitrogen gas (N2). These normally aerobic bacteria, mostly species of the genera Bacillus and Pseudomonas, use nitrate (NO3) as an electron acceptor in place of oxygen if they are kept under anaerobic conditions: 2 NO3 -+10 e- + 12 H+ → N2 + 6 H₂O
Nitrogen fixers convert atmospheric nitrogen gas into a chemical form usable by the nitrogen fixers themselves as well as by other organisms. They convert nitrogen gas to ammonia:
N2+6H>2 NH3
All organisms require nitrogen for their proteins, nucleic acids, and other important compounds. The vital process of nitrogen fixation is carried out by a wide variety of archaea and bacteria, including cyanobacteria, but by no other organisms. Ammonia is oxidized to nitrate in the soil and in seawater by chemolithotrophic bacteria called nitrifiers. Bacteria of two genera, Nitrosomonas and Nitrosococcus, convert ammonia to nitrite ions (NO₂-), and Nitrobacter oxidizes nitrite to nitrate (NO3).
What do the nitrifiers get out of these reactions? Their chemosynthesis is powered by the energy released by the oxidation of ammonia or nitrite. For example, by passing the electrons from nitrite through an electron transport chain, Nitrobacter can make ATP, and using
some of this ATP, it can also make NADH. With this ATP and NADH, the bacterium can convert CO2 and H2O to glucose.
Numerous bacteria base their metabolism on the modification of sulfur-containing ions and compounds in their environments. As examples, we have already mentioned the photoautotrophic bacteria and chemolithotrophic archaea that use H2S as an electron donor in place of H2O. Such uses of nitrogen and sulfur have environmental implications, as we’ll see in the next section.
Prokaryotes in Their Environments
Prokaryotes live in and exploit all sorts of environments and are part of all ecosystems. In the following pages, we’ll examine the roles of prokaryotes that live in soils, in water, and even in other living organisms, where they may exist in a neutral, benevolent, or parasitic
relationship with their host’s tissues.
Prokaryotes are important players in element cycling
Animals depend on photosynthetic plants and microorganisms for their food, directly or indirectly. But plants depend on other organisms-prokaryotes-for their own nutrition. The extent and diversity of life on Earth would not be possible without nitrogen fixation by prokaryotes. Nitrifiers are crucial to the biosphere because they convert the products of nitrogen fixation into nitrate ions, the form of nitrogen most easily used by many plants. Plants, in turn, are the source of nitrogen compounds for animals and fungi. Denitrifiers also play a key role in keeping the nitrogen cycle going. Without denitrifiers, which convert nitrate ions back into nitrogen gas, all forms of nitrogen would leach from the soil and end up in lakes and oceans, making life on land impossible. Other prokaryotes contribute to a similar cycle of sulfur. Prokaryotes, along with fungi, return tremendous quantities of organic carbon to the atmosphere as carbon dioxide.
In the ancient past, the cyanobacteria had an equally dramatic effect on life: Their photosynthesis generated oxygen, converting Earth from an anaerobic to an aerobic environment. The result was the wholesale loss of obligate anaerobic species that could not tolerate the O2 generated by the cyanobacteria. Only those anaerobes that were able to colonize environments that remained anaerobic survived. However, this transformation to aerobic environments made possible the evolution of cellular respiration and the subsequent explosion of
eukaryotic life. What other roles do prokaryotes play in the biosphere?